하플로그룹 R1b (Y-DNA)
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
R1b (Y-DNA)는 Y 염색체 상의 SNP(단일 염기 다형성) M343 변이로 정의되는 하플로그룹으로, 아프리카 외부의 남성 계통에서 가장 흔하게 나타난다. 약 18,500년 전에 시베리아와 중앙아시아에서 기원한 것으로 추정되며, 유럽, 아시아, 아프리카, 아메리카 등 전 세계적으로 분포한다. 특히 서유럽과 남유럽에서 높은 빈도를 보이며, 켈트계 민족과 바스크인에게서 80% 이상으로 나타난다. R1b는 인도유럽어족의 확산과 밀접한 관련이 있으며, 얌나 문화와 관련된 고대 DNA에서 높은 빈도로 발견된다. R1b는 켄툼어군 언어와 연관되며, 청동기 시대에 유럽으로 확산되었다. R1b의 하위 그룹인 R-V88은 아프리카 사헬 지역에서 주로 발견되며, 아프리카로의 역이동을 보여주는 증거로 해석된다.
R1b는 약 18,500년 전에 발생한 것으로 추정되며, 초기에는 시베리아 및 중앙아시아 지역에 분포했을 것으로 보인다. 이는 타티아나 카라페트 외 (2008)의 연구에 따른 것이며, R1b는 R1이 출현한 직후에 발생했을 것이다.[60]
2. 기원 및 확산
R1b를 보유한 것으로 밝혀진 초기 인류 유해는 다음과 같다.
R1b는 "매크로-하플로그룹" K (M9) 내의 하위 그룹이며, 아프리카 외부의 인간 남성 계통에서 가장 흔한 그룹이다. K는 동남아시아에서 기원한 것으로 여겨지며, 카라페트 T. 외 (2014)는 "K-M526의 급격한 다양화 과정이 동남아시아에서 발생했을 가능성이 높고, 그 후 R과 Q의 조상이 서쪽으로 확장되었다"고 제안했다.[7] 그러나 R*의 가장 오래된 예는 고대 북유라시아인 샘플(24,000년 전의 말타 소년)에서 발견되었으며, 그 전구체인 P1은 약 31,600년 전의 북부 시베리아의 또 다른 고대 북유라시아인 샘플(야나 RHS)에서 발견되었다.[8]
마리야 김부타스의 쿠르간 가설에 따르면, 현재 유럽에서 가장 흔한 하플로그룹 R1b-M269와 R1a는 인도유럽어족과 함께 서유라시아 스텝에서 확장되었을 것이며, 현대 유럽인에게는 존재하지만 신석기 유럽인에게는 존재하지 않았던 상염색체 성분이 R1b 및 R1a의 부계 계통 및 인도유럽어와 함께 도입되었을 것으로 추정했다.[1][2]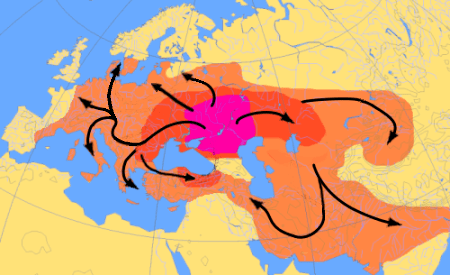
초기 신석기 시대 중앙 및 북부 유럽의 선형 도기 문화 정착지 유적에서는 하플로그룹 R1b-M269에 속하는 남성이 아직 발견되지 않았다.[9][10] 서유럽, 특히 영국에서 하플로그룹 R1b-M269의 확산은 비커 문화의 확산으로 추적되며, 청동기 시대 초인 약 5,000–4,500년 전에 서유럽에서 R1b-M269 하플로그룹이 갑자기 나타났다.[11]
=== R1b의 초기 분기 ===
R1b* (R-M343*)의 확인된 사례는 동료 심사를 거친 문헌에서 보고된 적이 없다.[12] 초기 연구에서는 R1b의 가장 흔한 형태인 R-M269, R-M73, R-V88을 제외한 R1b (xM73, xM269)의 예시를 "R1b*"의 기저 사례로 추정하기도 했다.[12] 그러나 R-M343 (xM73, M269, V88)은 드물지만, R-L278* (R1b*), R-L389* (R1b1a*), R-P297* (R1b1a1*), R-V1636 (R1b1a2) 또는 R-PH155 (R1b2)와 같은 R1b의 드물고 나중에 발견된 하위 분기에 속할 가능성을 배제할 수 없다.[19]
R-M343 (xM73, M269, V88) 비율이 가장 높은 인구는 카자흐스탄 남동부의 쿠르드족 (13%)으로 알려져 있었으나,[56] 최근 이란의 Y 염색체 변이 연구에서는 이란 하위 집단에서 R-M343 (xV88, M73, M269)이 4.3%로 나타났다.[18] 이러한 사례 중 일부는 R1b* (R-M343*), R1a* (R-M420*), R1 (R-M173)의 문서화되지 않은 분기이거나, 양성 상태에서 음성 조상 상태로의 마커 역 돌연변이일 가능성도 있다.[19]
R1b의 분포에 관한 이전 연구 편집 내용은 Cruciani et al. (2010)에서 찾을 수 있다.[21]
| 대륙 | 지역 | 표본 크기 | 총 R1b | R-P25 (신뢰할 수 없는 마커 R1b1*) | R-V88 (R1b1b) | R-M269 (R1b1a1a2) | R-M73 (R1b1a1a1) |
|---|---|---|---|---|---|---|---|
| 아프리카 | 북아프리카 | 691 | 5.9% | 0.0% | 5.2% | 0.7% | 0.0% |
| 아프리카 | 중앙 사헬 지역 | 461 | 23.0% | 0.0% | 23.0% | 0.0% | 0.0% |
| 아프리카 | 서아프리카 | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 아프리카 | 동아프리카 | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 아프리카 | 남아프리카 | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 유럽 | 서유럽 | 465 | 57.8% | 0.0% | 0.0% | 57.8% | 0.0% |
| 유럽 | 북서유럽 | 43 | 55.8% | 0.0% | 0.0% | 55.8% | 0.0% |
| 유럽 | 중앙유럽 | 77 | 42.9% | 0.0% | 0.0% | 42.9% | 0.0% |
| 유럽 | 북동유럽 | 74 | 1.4% | 0.0% | 0.0% | 1.4% | 0.0% |
| 유럽 | 러시아인 | 60 | 6.7% | 0.0% | 0.0% | 6.7% | 0.0% |
| 유럽 | 동유럽 | 149 | 20.8% | 0.0% | 0.0% | 20.8% | 0.0% |
| 유럽 | 동남유럽 | 510 | 13.1% | 0.0% | 0.2% | 12.9% | 0.0% |
| 아시아 | 서아시아 | 328 | 5.8% | 0.0% | 0.3% | 5.5% | 0.0% |
| 아시아 | 남아시아 | 288 | 4.8% | 0.0% | 0.0% | 1.7% | 3.1% |
| 아시아 | 동남아시아 | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 아시아 | 북동아시아 | 30 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 아시아 | 동아시아 | 156 | 0.6% | 0.0% | 0.0% | 0.6% | 0.0% |
| 합계 | 5326 | ||||||
R-L278은 현대 남성에서 R-L754와 R-PH155 하위 그룹으로 분류되지만, R-L278*가 존재할 가능성도 있다. 고대 DNA에서도 사례가 존재할 수 있지만, 품질 문제로 하위 그룹 확인이 어려운 경우가 많다. 오래된 기사에서 설명된 일부 사례는 R1b1b (R-V88)에 속하는 것으로 추정된다.[20] 대부분의 R1b 사례는 R1b1b (R-V88) 또는 R1b1a (R-P297)에 속하며, Cruciani et al. (2010)의 연구에서는 이탈리아인 3건, 서아시아인 1건, 동아시아인 1건이 발견되었다.[21] 우크라이나 드네스트르강–카르파티아 산맥 지역 연구에서는 P25 양성, M269 음성인 3건이 발견되었다.[22]
R-L754는 R1b의 대다수를 포함한다. R-L754*(xL389, V88)의 유일하게 알려진 예는 에피그라베티안 문화에 속했던 빌라브루나 1 (기원전 14,000년경, 이탈리아 북동부)이다.
=== 얌나 문화와 R1b-M269의 확산 ===
R-M269는 현재 유럽 남성에게서 가장 흔한 Y-DNA 계통으로, 유럽 남성 약 1억 1천만 명이 이 유전자를 가지고 있는 것으로 추정된다.[33] R-M269는 인도유럽어족의 확장과의 관련성 때문에 과학적 및 대중적 관심이 높다.[32] 특히 R-Z2103 하위 분류는 얌나 문화와 관련된 고대 DNA에서 널리 발견되었다. 얌나 문화 집단의 7명 모두가 R1b-M269 하위 분류에 속하는 것으로 밝혀졌다.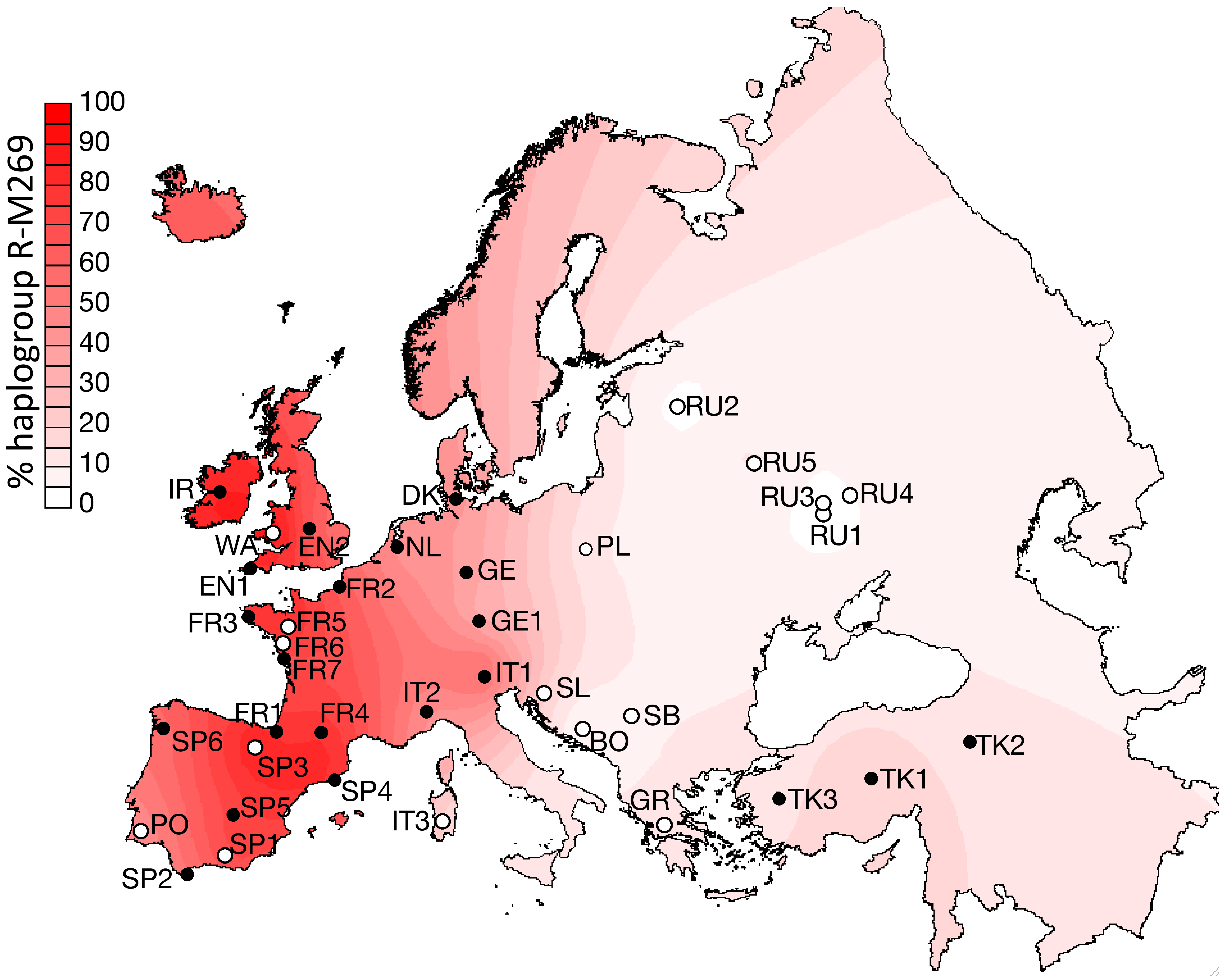
이전 연구에서는 R-M269가 서아시아에서 기원하여 신석기 시대에 유럽에 존재했을 가능성이 높다고 제안했다.[13][56][34][35] 그러나 고대 유물의 DNA 연구 결과, 청동기 시대 이전에 유럽에서 R-M269가 부족했고, 유럽 내 하위 분류의 분포는 청동기 시대와 철기 시대의 다양한 이주에 크게 기인한다는 것이 밝혀졌다. R-M269에 속하는 것으로 분류된 가장 오래된 샘플은 서아시아가 아닌 동유럽과 폰토스-카스피 스텝에서 발견되었다. 서유럽 인구는 R-M412 (R-L51)의 R-P312/S116 및 R-U106/S21 하위 분류로 나뉜다.
R-M269의 유럽 내 분포는 동쪽에서 서쪽으로 갈수록 빈도가 증가한다. 웨일스에서 92%, 아일랜드에서 82%, 스코틀랜드에서 70%, 스페인에서 68%, 프랑스에서 60% (노르망디에서는 76%), 포르투갈에서 약 60%,[56] 독일에서 50%, 네덜란드에서 50%, 이탈리아에서 47%,[36] 동부 잉글랜드에서 45% 그리고 아이슬란드에서 42%로 최고치를 기록한다. R-M269는 아일랜드 일부 지역에서 95%에 달하는 수준에 이른다.
R-M269는 중앙 유라시아 전역에서 낮은 빈도로 발견되었지만,[3] 페름 지역의 바시키르인에서는 비교적 높은 빈도(84.0%)를 보였다.[37] 이 마커는 중국과 인도에서 1% 미만의 빈도로 나타난다. 북아프리카와 인접한 섬에서는 R-V88 (R1b1b)이 더 강하게 나타나지만, R-M269는 고대부터 존재했던 것으로 보인다. 예를 들어, 카나리아 제도의 푼타 아줄에서 11세기에서 13세기에 해당하는 유적에서 약 44%의 비율로 R-M269가 발견되었다. 이 유적은 빔바체 (또는 비마페)와 관련이 있는데, 이들은 관체족의 하위 집단이다.[38] 살아있는 남성에서는 특히 알제리를 비롯한 북아프리카 일부 지역에서 10%의 비율로 최고치를 기록한다.[39] 사하라 사막 이남 아프리카에서는 나미비아에서 헤레로 남성 중 8%의 비율로 R-M269가 최고치를 보이는 것으로 나타났다.[58] 서아시아에서는 아르메니아인 남성의 40%, 투르크멘인 남성의 35% 이상에서 R-M269가 보고되었다.[40][41]
분화되지 않은 기저 R-M269* 외에, R-M269에는 두 개의 주요 분기가 있다.
- R-L23 (R1b1a1b1; L23/PF6534/S141) 및
- R-PF7558 (R1b1a1b2; PF7558/PF7562.)
R-L23 (Z2105/Z2103)은 이델-우랄의 사람들에게서 보고되었다.[42] 파라그룹 R-M269(xL23) 내의 하위 분류는 발칸 반도에서 가장 높은 빈도로 발견되는 것으로 보인다.[56] 코소보, 폴란드 및 남동부 바시키르 공화국의 바시키르인은 R-L23 (xM412)의 높은 비율을 가지고 있다는 점에서 주목할 만하다.[56] 아라라트 평원의 아르메니아에서 검사한 표본에서 일부는 R-M269(xL23)에 속했고 일부는 R-L23*에 속했으며, L23의 알려진 하위 분류에 속하는 사람은 없었다.[43]
2009년, 서기 670년경으로 추정되는 에르골딩 (바이에른, 독일)의 초기 중세 매장지에서 추출한 DNA의 결과, 일부는 하플로그룹 R1b로 밝혀졌고, 일부는 하플로그룹 G2a에 속했다.[44]
=== 아프리카로의 역이동 (R-V88) ===
R1b1b (PF6279/V88)는 SNP 마커 V88의 존재로 정의되며, 2010년 Cruciani et al.에 의해 발견이 발표되었다.[21] 남부 유럽과 서아시아를 제외하면, R-V88의 대부분은 사헬 지역, 특히 아프리카아시아어족의 차드어족 분파를 사용하는 집단에서 발견된다.
D'Atanasio et al. (2018)은 R1b-V88이 약 12,000년 전 유럽에서 기원하여 '녹색 사하라' 기간인 8,000~7,000년 전 북아프리카로 이동했다고 제안했다.[45] R1b-V88 내 주요 하위 그룹인 R1b-V1589는 약 5,500년 전 차드호 분지 지역에서 확장되었고, 일부 계통은 사하라 사막을 넘어 북아프리카로 다시 이동했다.[45]
Marcus et al. (2020)은 북쪽에서 남쪽으로의 사하라 사막 횡단 이동에 대한 강력한 증거를 제시한다. 가장 초기의 기저 R1b-V88 하플로그룹은 약 11,000년 전 여러 동유럽 수렵 채집인에게서 발견된다.[46] 이 하플로그룹은 신석기 시대 농부의 확장과 함께 퍼진 것으로 보이며, 이들은 약 7,500년 전에 서부 지중해에 농업을 정착시켰다. R1b-V88 하플로그룹은 고대 신석기 시대 독일, 이탈리아 중부, 이베리아 반도, 특히 사르데냐에서 높은 빈도로 확인되었다.[46][47][48] 오늘날 아프리카 하플로그룹(V2197)으로 이어지는 분파의 일부는 이미 스페인과 사르데냐의 신석기 시대 유럽 개인에게서 파생되었다.[46][47][48] 유당 분해 효소 유지 대립 유전자 또한 풀라니족(Fulani)과 투부족(Toubou)과 같이 R1b-V88을 높은 빈도로 가진 아프리카 집단에서 확인되었다.[49][50][47][51] 유럽 신석기 시대 농부가 아프리카에 존재한다는 것은 기원전 5400년경부터의 모로코 샘플로 더욱 입증되었다.[52][53]
요르단, 이집트, 수단에서 높은 수준의 "R1b*"가 보고되었으나,[54][58][55] Myres et al. (2011)의 후속 연구는 해당 샘플이 대부분 하위 그룹 R-V88에 속할 가능성이 있음을 나타낸다.[56] Myres et al. (2011)에 따르면, 이는 R1b를 가진 사람들이 아시아에서 아프리카로 역이동한 것으로 설명될 수 있다.[56]
R-V88은 서부 중앙아프리카의 북부 카메룬에서 매우 높은 빈도로 발견되며, 이는 유라시아에서 온 사람들의 이슬람 이전의 이동으로 인한 것으로 여겨진다.[58][59] R-V88의 두 분파인 R-M18과 R-V35는 거의 사르데냐 섬에서만 발견된다.
다른 연구와는 달리, Shriner & Rotimi (2018)는 R1b가 차드에 유입된 것을 바가라 아랍인의 최근 이동과 연관시켰다.[57]
2. 1. R1b의 초기 분기
R1b* (R-M343*)의 확인된 사례는 동료 심사를 거친 문헌에서 보고된 적이 없다.[12] 초기 연구에서는 R1b의 가장 흔한 형태인 R-M269, R-M73, R-V88을 제외한 R1b (xM73, xM269)의 예시를 "R1b*"의 기저 사례로 추정하기도 했다.[12] 그러나 파라그룹 R-M343 (xM73, M269, V88)은 드물지만, R-L278* (R1b*), R-L389* (R1b1a*), R-P297* (R1b1a1*), R-V1636 (R1b1a2) 또는 R-PH155 (R1b2)와 같은 R1b의 드물고 나중에 발견된 하위 분기에 속할 가능성을 배제할 수 없다.[19]R-M343 (xM73, M269, V88) 비율이 가장 높은 인구는 카자흐스탄 남동부의 쿠르드족 (13%)으로 알려져 있었으나,[56] 최근 이란의 Y 염색체 변이 연구에서는 이란 하위 집단에서 R-M343 (xV88, M73, M269)이 4.3%로 나타났다.[18] 이러한 사례 중 일부는 R1b* (R-M343*), R1a* (R-M420*), R1 (R-M173)의 문서화되지 않은 분기이거나, 양성 상태에서 음성 조상 상태로의 마커 역 돌연변이일 가능성도 있다.[19]
R1b의 분포에 관한 이전 연구 편집 내용은 Cruciani et al. (2010)에서 찾을 수 있다.[21]
| 대륙 | 지역 | 표본 크기 | 총 R1b | R-P25 (신뢰할 수 없는 마커 R1b1*) | R-V88 (R1b1b) | R-M269 (R1b1a1a2) | R-M73 (R1b1a1a1) |
|---|---|---|---|---|---|---|---|
| 아프리카 | 북아프리카 | 691 | 5.9% | 0.0% | 5.2% | 0.7% | 0.0% |
| 아프리카 | 중앙 사헬 지역 | 461 | 23.0% | 0.0% | 23.0% | 0.0% | 0.0% |
| 아프리카 | 서아프리카 | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 아프리카 | 동아프리카 | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 아프리카 | 남아프리카 | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 유럽 | 서유럽 | 465 | 57.8% | 0.0% | 0.0% | 57.8% | 0.0% |
| 유럽 | 북서유럽 | 43 | 55.8% | 0.0% | 0.0% | 55.8% | 0.0% |
| 유럽 | 중앙유럽 | 77 | 42.9% | 0.0% | 0.0% | 42.9% | 0.0% |
| 유럽 | 북동유럽 | 74 | 1.4% | 0.0% | 0.0% | 1.4% | 0.0% |
| 유럽 | 러시아인 | 60 | 6.7% | 0.0% | 0.0% | 6.7% | 0.0% |
| 유럽 | 동유럽 | 149 | 20.8% | 0.0% | 0.0% | 20.8% | 0.0% |
| 유럽 | 동남유럽 | 510 | 13.1% | 0.0% | 0.2% | 12.9% | 0.0% |
| 아시아 | 서아시아 | 328 | 5.8% | 0.0% | 0.3% | 5.5% | 0.0% |
| 아시아 | 남아시아 | 288 | 4.8% | 0.0% | 0.0% | 1.7% | 3.1% |
| 아시아 | 동남아시아 | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 아시아 | 북동아시아 | 30 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| 아시아 | 동아시아 | 156 | 0.6% | 0.0% | 0.0% | 0.6% | 0.0% |
| 합계 | 5326 | ||||||
R-L278은 현대 남성에서 R-L754와 R-PH155 하위 그룹으로 분류되지만, R-L278*가 존재할 가능성도 있다. 고대 DNA에서도 사례가 존재할 수 있지만, 품질 문제로 하위 그룹 확인이 어려운 경우가 많다. 오래된 기사에서 설명된 일부 사례는 R1b1b (R-V88)에 속하는 것으로 추정된다.[20] 대부분의 R1b 사례는 R1b1b (R-V88) 또는 R1b1a (R-P297)에 속하며, Cruciani et al. (2010)의 연구에서는 이탈리아인 3건, 서아시아인 1건, 동아시아인 1건이 발견되었다.[21] 우크라이나 드네스트르강–카르파티아 산맥 지역 연구에서는 P25 양성, M269 음성인 3건이 발견되었다.[22]
R-L754는 R1b의 대다수를 포함한다. R-L754*(xL389, V88)의 유일하게 알려진 예는 에피그라베티안 문화에 속했던 빌라브루나 1 (기원전 14,000년경, 이탈리아 북동부)이다.
2. 2. 얌나 문화와 R1b-M269의 확산
R-M269는 현재 유럽 남성에게서 가장 흔한 Y-DNA 계통으로, 유럽 남성 약 1억 1천만 명이 이 유전자를 가지고 있는 것으로 추정된다.[33] R-M269는 인도유럽어족의 확장과의 관련성 때문에 과학적 및 대중적 관심이 높다.[32] 특히 R-Z2103 하위 분류는 얌나 문화와 관련된 고대 DNA에서 널리 발견되었다. 얌나 문화 집단의 7명 모두가 R1b-M269 하위 분류에 속하는 것으로 밝혀졌다.이전 연구에서는 R-M269가 서아시아에서 기원하여 신석기 시대에 유럽에 존재했을 가능성이 높다고 제안했다.[13][56][34][35] 그러나 고대 유물의 DNA 연구 결과, 청동기 시대 이전에 유럽에서 R-M269가 부족했고, 유럽 내 하위 분류의 분포는 청동기 시대와 철기 시대의 다양한 이주에 크게 기인한다는 것이 밝혀졌다. R-M269에 속하는 것으로 분류된 가장 오래된 샘플은 서아시아가 아닌 동유럽과 폰토스-카스피 스텝에서 발견되었다. 서유럽 인구는 R-M412 (R-L51)의 R-P312/S116 및 R-U106/S21 하위 분류로 나뉜다.
R-M269의 유럽 내 분포는 동쪽에서 서쪽으로 갈수록 빈도가 증가한다. 웨일스에서 92%, 아일랜드에서 82%, 스코틀랜드에서 70%, 스페인에서 68%, 프랑스에서 60% (노르망디에서는 76%), 포르투갈에서 약 60%,[56] 독일에서 50%, 네덜란드에서 50%, 이탈리아에서 47%,[36] 동부 잉글랜드에서 45% 그리고 아이슬란드에서 42%로 최고치를 기록한다. R-M269는 아일랜드 일부 지역에서 95%에 달하는 수준에 이른다.
R-M269는 중앙 유라시아 전역에서 낮은 빈도로 발견되었지만,[3] 페름 지역의 바시키르인에서는 비교적 높은 빈도(84.0%)를 보였다.[37] 이 마커는 중국과 인도에서 1% 미만의 빈도로 나타난다. 북아프리카와 인접한 섬에서는 R-V88 (R1b1b)이 더 강하게 나타나지만, R-M269는 고대부터 존재했던 것으로 보인다. 예를 들어, 카나리아 제도의 푼타 아줄에서 11세기에서 13세기에 해당하는 유적에서 약 44%의 비율로 R-M269가 발견되었다. 이 유적은 빔바체 (또는 비마페)와 관련이 있는데, 이들은 관체족의 하위 집단이다.[38] 살아있는 남성에서는 특히 알제리를 비롯한 북아프리카 일부 지역에서 10%의 비율로 최고치를 기록한다.[39] 사하라 사막 이남 아프리카에서는 나미비아에서 헤레로 남성 중 8%의 비율로 R-M269가 최고치를 보이는 것으로 나타났다.[58] 서아시아에서는 아르메니아인 남성의 40%, 투르크멘인 남성의 35% 이상에서 R-M269가 보고되었다.[40][41]
분화되지 않은 기저 R-M269* 외에, R-M269에는 두 개의 주요 분기가 있다.
- R-L23 (R1b1a1b1; L23/PF6534/S141) 및
- R-PF7558 (R1b1a1b2; PF7558/PF7562.)
R-L23 (Z2105/Z2103)은 이델-우랄의 사람들에게서 보고되었다.[42] 파라그룹 R-M269(xL23) 내의 하위 분류는 발칸 반도에서 가장 높은 빈도로 발견되는 것으로 보인다.[56] 코소보, 폴란드 및 남동부 바시키르 공화국의 바시키르인은 R-L23 (xM412)의 높은 비율을 가지고 있다는 점에서 주목할 만하다.[56] 아라라트 평원의 아르메니아에서 검사한 표본에서 일부는 R-M269(xL23)에 속했고 일부는 R-L23*에 속했으며, L23의 알려진 하위 분류에 속하는 사람은 없었다.[43]
2009년, 서기 670년경으로 추정되는 에르골딩 (바이에른, 독일)의 초기 중세 매장지에서 추출한 DNA의 결과, 일부는 하플로그룹 R1b로 밝혀졌고, 일부는 하플로그룹 G2a에 속했다.[44]
2. 3. 아프리카로의 역이동 (R-V88)
R1b1b (PF6279/V88)는 SNP 마커 V88의 존재로 정의되며, 2010년 Cruciani et al.에 의해 발견이 발표되었다.[21] 남부 유럽과 서아시아를 제외하면, R-V88의 대부분은 사헬 지역, 특히 아프리카아시아어족의 차드어족 분파를 사용하는 집단에서 발견된다.D'Atanasio et al. (2018)은 R1b-V88이 약 12,000년 전 유럽에서 기원하여 '녹색 사하라' 기간인 8,000~7,000년 전 북아프리카로 이동했다고 제안했다.[45] R1b-V88 내 주요 하위 그룹인 R1b-V1589는 약 5,500년 전 차드호 분지 지역에서 확장되었고, 일부 계통은 사하라 사막을 넘어 북아프리카로 다시 이동했다.[45]
Marcus et al. (2020)은 북쪽에서 남쪽으로의 사하라 사막 횡단 이동에 대한 강력한 증거를 제시한다. 가장 초기의 기저 R1b-V88 하플로그룹은 약 11,000년 전 여러 동유럽 수렵 채집인에게서 발견된다.[46] 이 하플로그룹은 신석기 시대 농부의 확장과 함께 퍼진 것으로 보이며, 이들은 약 7,500년 전에 서부 지중해에 농업을 정착시켰다. R1b-V88 하플로그룹은 고대 신석기 시대 독일, 이탈리아 중부, 이베리아 반도, 특히 사르데냐에서 높은 빈도로 확인되었다.[46][47][48] 오늘날 아프리카 하플로그룹(V2197)으로 이어지는 분파의 일부는 이미 스페인과 사르데냐의 신석기 시대 유럽 개인에게서 파생되었다.[46][47][48] 유당 분해 효소 유지 대립 유전자 또한 풀라니족(Fulani)과 투부족(Toubou)과 같이 R1b-V88을 높은 빈도로 가진 아프리카 집단에서 확인되었다.[49][50][47][51] 유럽 신석기 시대 농부가 아프리카에 존재한다는 것은 기원전 5400년경부터의 모로코 샘플로 더욱 입증되었다.[52][53]
요르단, 이집트, 수단에서 높은 수준의 "R1b*"가 보고되었으나,[54][58][55] Myres et al. (2011)의 후속 연구는 해당 샘플이 대부분 하위 그룹 R-V88에 속할 가능성이 있음을 나타낸다.[56] Myres et al. (2011)에 따르면, 이는 R1b를 가진 사람들이 아시아에서 아프리카로 역이동한 것으로 설명될 수 있다.[56]
R-V88은 서부 중앙 아프리카의 북부 카메룬에서 매우 높은 빈도로 발견되며, 이는 유라시아에서 온 사람들의 이슬람 이전의 이동으로 인한 것으로 여겨진다.[58][59] R-V88의 두 분파인 R-M18과 R-V35는 거의 사르데냐 섬에서만 발견된다.
다른 연구와는 달리, Shriner & Rotimi (2018)는 R1b가 차드에 유입된 것을 바가라 아랍인의 최근 이동과 연관시켰다.[57]
2. 4. 동아시아의 R1b
3. 분포
하플로그룹 R1b (Y-DNA) 계통은 서유럽과 남유럽에 현저하게 분포하는 계통이며, 바스크인과 켈트계 민족에게 80% 이상의 높은 빈도로 나타난다.[87][88][89][90]
얌나 문화인에게서 높은 빈도로 발견되는[91][92] 것으로부터, 인도유럽어족의 담당자이다. (R1b는 켄툼어군, R1a는 사템어군의 담당자이다[107]) 유럽 서부에는 청동기 시대에 분포를 넓혔으며[93], 우네티체 문화의 담당자[94]로 여겨진다.
또한 토하라어족과 아파나시에보 문화의 담당자이다.[95]
대항해 시대 이후의 인종 이동에 따라, 남북 아메리카 대륙과 오스트레일리아 대륙에도 분포 범위를 확대했다.
또한 바시키르인에게서도 86%의 높은 빈도로 나타난다.[96]
이 흐름과는 별개로, 고대에도 아프리카 내륙부로 이동한 R1b 계통의 한 집단이 있었던 것으로 보이며, 하플로그룹 R-V88로 정의되는 분기 계통이 차드어족의 언어를 사용하는 카메룬의 원주민 사이에서 고농도로 발견되고 있다.[97][98][99]
3. 1. 유럽
R-M269는 현재 유럽 남성에게 가장 흔한 Y-DNA 계통으로, 유럽 남성 약 1억 1천만 명이 이 유전자를 가지고 있는 것으로 추정된다.[33] R-M269의 유럽 내 분포는 동쪽에서 서쪽으로 갈수록 빈도가 증가한다. 웨일스에서 92%, 아일랜드에서 82%, 스코틀랜드에서 70%, 스페인에서 68%, 프랑스에서 60% (노르망디에서는 76%), 포르투갈에서 약 60%,[56] 독일에서 50%, 네덜란드에서 50%, 이탈리아에서 47%,[36] 동부 잉글랜드에서 45% 그리고 아이슬란드에서 42%로 최고치를 기록한다. 특히 아일랜드 일부 지역에서는 R-M269가 95%에 달하는 수준에 이른다. 바스크인과 켈트계 민족에게 80% 이상의 높은 빈도로 나타난다.[87][88][89][90]R-M269는 인도유럽어족의 확장과의 관련성 때문에 과학적 및 대중적 관심이 높다. 특히 R-Z2103 하위 분류는 얌나 문화와 관련된 고대 DNA에서 널리 발견되었다.[91][92] R1b는 켄툼어군과 관련이 있으며, 청동기 시대에 유럽 서부에 분포를 넓혔고,[93] 우네티체 문화의 담당자로 여겨진다.[94]
대항해 시대 이후의 인종 이동에 따라, 남북 아메리카 대륙과 오스트레일리아 대륙에도 분포 범위를 확대했다.
분화되지 않은 기저 R-M269* 외에, (2017년 기준) R-M269에는 R-L23 (R1b1a1b1; L23/PF6534/S141)와 R-PF7558 (R1b1a1b2; PF7558/PF7562)의 두 가지 주요 분기가 있다. 파라그룹 R-M269(xL23) 내의 하위 분류는 중앙 발칸 반도에서 가장 높은 빈도로 발견되는 것으로 보이는데, 특히 코소보에서 7.9%, 북마케도니아에서 5.1%, 세르비아에서 4.4%로 나타난다.[56]
2009년, 서기 670년경으로 추정되는 에르골딩 (바이에른, 독일)의 초기 중세 매장지에서 6구의 뼈에서 추출한 DNA를 분석한 결과, 4구는 하플로그룹 G2a에 속했고, 나머지 2구는 독일, 아일랜드, 미국의 현대 인구와 가장 가까운 하플로그룹 R1b로 밝혀졌다.[44]
3. 2. 아시아
R1b는 중앙아시아에서도 발견되며, 특히 바시키르인에게서 높은 빈도로 나타난다.[56] 남아시아와 동아시아에서는 낮은 빈도로 나타난다. 서아시아에서는 아르메니아인과 투르크멘인에게서 비교적 높은 빈도로 나타난다.Malyarchuk 외. (2011)의 연구에 따르면 쇼르족의 13.2% (5/38), 텔레우트족의 11.4% (5/44)에서 R-M73이 발견되었다.[23] Dulik 외. (2012)는 러시아 알타이 공화국의 쿠만딘 샘플의 35.3% (6/17)에서 R-M73을 발견했다.[24] 이들 중 일부는 칼미크족, 투바족, 알타이족의 Y-STR 핵형과 유사한 핵형을 공유한다.[23]
Myres 외. (2011)는 바슈코르토스탄 남동부의 바시키르족 샘플에서 높은 빈도의 R-M73 (77/329 = 23.4% R1b-M73)을 발견했다.[56] 이외에도 발카르족(10.3%), 파키스탄 북부(9.4%), 카라차이족(5.8%) 등에서도 R-M73이 발견되었다.[56]
Di Cristofaro 외. (2013)은 중앙 몽골의 몽골족의 11.1% (2/18), 키르기스스탄 남서부의 키르기스족의 5.0% (1/20) 등에서 R-M478/M73을 발견했다.[27] 이들 중 일부는 Dulik 외. (2012)가 연구한 R-M73의 쿠만딘 구성원과 동일한 Y-STR 핵형 클러스터에 속하며, 이 클러스터의 가장 두드러진 Y-STR 값은 DYS390=19이다.[56]
Karafet 외. (2018)은 텔레우트족 샘플의 37.5% (15/40), 신장 위구르 자치구의 위구르족 샘플의 4.5% (3/66) 등에서 R-M73을 발견했다.[29] 이들 중 일부는 쿠만딘-몽골 R-M73 클러스터에 속하는 것으로 보인다.
2017년 카자흐스탄 연구에서는 카자흐스탄의 카자흐족 샘플의 3.17% (41/1294)에서 하플로그룹 R1b-M478 Y-DNA를 발견했으며, 특정 카자흐 부족 구성원들 사이에서 평균보다 높은 빈도로 관찰되었다.[30] 2018년 중국 연구에서는 신장 아와트 현 호리콜향의 돌란족 위구르족 샘플의 9.2% (7/76)에서 하플로그룹 R1b-M478 Y-DNA를 발견했다.[31]
3. 3. 아프리카
R1b는 북아프리카 일부 지역에서 10% 정도의 비율로 나타나며, 특히 알제리의 모자비테 베르베르족에게서 3.0%[58][59], 이집트 시와 베르베르족에게서 28.0%[58][59]의 빈도를 보인다. R1b1b (PF6279/V88; 이전에는 R1b1a2)는 SNP 마커 V88의 존재에 의해 정의되며, 남부 유럽과 서아시아의 개인을 제외하고 R-V88의 대부분은 사헬 지역, 특히 아프리카아시아어족의 차드어족 분파를 사용하는 집단에서 발견되었다.[21]D'Atanasio et al. (2018)은 R1b-V88이 약 12,000년 전에 유럽에서 기원하여 '녹색 사하라' 기간 동안 8,000~7,000년 전에 북아프리카로 이동했다고 제안했다. R1b-V88 내의 주요 하위 그룹인 R1b-V1589는 약 5,500년 전에 차드호 분지 지역에서 추가 확장을 겪었고, 일부 계통은 사하라 사막을 넘어 북아프리카로 다시 이동했다.[45]
Marcus et al. (2020)은 북쪽에서 남쪽으로의 사하라 사막 횡단 이동에 대한 강력한 증거를 제시한다. 가장 초기의 기저 R1b-V88 하플로그룹은 약 11,000년 전에 여러 동유럽 수렵 채집인에서 발견된다. 이 하플로그룹은 신석기 시대 농부의 확장과 함께 퍼진 것으로 보이며, 이들은 약 7500 BP에 서부 지중해에 농업을 정착시켰다. R1b-V88 하플로그룹은 고대 신석기 시대의 독일, 이탈리아 중부, 이베리아 반도, 특히 사르데냐에서 높은 빈도로 확인되었다. 오늘날 아프리카 하플로그룹(V2197)으로 이어지는 분파의 일부는 이미 스페인과 사르데냐의 신석기 시대 유럽 개인에게서 파생되었으며, 이는 북쪽에서 남쪽으로의 사하라 사막 횡단 이동을 더욱 뒷받침한다.[46][47][48] 유럽의 자가 염색체 조상, mtDNA 하플로그룹, 그리고 유당 분해 효소 유지 대립 유전자 또한 풀라니족(Fulani)과 투부족(Toubou)과 같이 R1b-V88을 높은 빈도로 가진 아프리카 집단에서 확인되었다.[49][50][47][51] 유럽 신석기 시대 농부가 아프리카에 존재한다는 것은 기원전 5400년경부터의 모로코 샘플로 더욱 입증되었다.[52][53]
사하라 사막 이남 아프리카에서는 카메룬 북부의 차드어족 언어 사용자들에게서 매우 높은 빈도로 나타난다. 특히 울데메족(95.5%), 마다족(82.4%), 마파족(87.5%), 기지거족(77.8%), 기다르족(66.7%) 등에게서 높은 빈도를 보인다.[58][59] 이는 유라시아에서 온 사람들의 이슬람 이전의 이동으로 인한 것으로 여겨진다.[58][59] 반면, Shriner & Rotimi (2018)는 R1b가 차드에 유입된 것을 바가라 아랍인의 최근 이동과 연관시켰다.[57]
3. 4. 아메리카 및 오세아니아
대항해 시대 이후 유럽인의 이주로 인해 남북 아메리카 대륙과 오스트레일리아 대륙에도 R1b가 분포하게 되었다.[87][88][89][90]4. 특징 및 의의
하플로그룹 R1b는 인도유럽어족의 확산과 밀접한 관련이 있는 것으로 추정된다.[102][103] 특히 R-M269 하위 분파는 얌나 문화와 관련이 깊다.[104] 얌나 문화의 유골에서는 하플로그룹 R1b (Y 염색체)가 91.5%의 높은 빈도로 검출되었지만, 하플로그룹 R1a는 검출되지 않았다.[104]
바스크인은 비인도유럽어족 언어를 사용하지만, R1b의 빈도가 매우 높아 유전적 부동 또는 언어 교체 현상과 관련이 있을 것으로 추정된다.[107] 일설에는 인도유럽어를 사용하는 남성 정복자 집단이 원주 바스크인 여성에게 많은 자녀를 낳았지만, 그 아이들은 아버지의 인도유럽어가 아닌 어머니의 바스크어로 양육되었기 때문에, 부계만 R1b의 인도유럽어계이면서 언어는 바스크어를 유지했다는 설명도 있다.[107]
R1b는 적발 유전자와 상관관계가 있는 것으로 알려져 있다.[105]
이집트 제18왕조의 투탕카멘 미라의 Y-DNA 하플로그룹은 R-M269로 확인되었는데,[68][69] 이는 유럽에 분포하는 유형으로, 현재 이집트인에게는 거의 나타나지 않는다.[108][109]
KV55에서 발견된 미라는 투탕카멘의 아버지로 추정되며, 아멘호테프 3세로 부터 R1b를 물려받았다.[68][69]
스피티흐네프 1세의 유해도 R1b로 나타났고[66], 부르봉 왕가는 R1b1b 하플로그룹을 가지고 있다.[67]
북아메리카 원주민 중 알곤킨계 민족에게서 보이는 하플로그룹 R1은 대부분 R1b이다. 11세기 무렵, 바스크인 어부들이 뉴펀들랜드 해안, 세인트로렌스 강 하구에 이르렀다는 기록이 있으며, 16세기에 자크 카르티에의 탐험대가 세인트로렌스 강을 발견했을 때, 그곳에서는 바스크인 어선단이 어업을 하고 있었다고 한다.[110]
5. 한국의 R1b
6. 계통수
R1b (M343)는 SNP 돌연변이 M343으로 정의되는 Y 염색체 하플로그룹이다.[20] R1b, R1b1 등과 같은 이름은 계통 발생학적 이름으로, 하플로그룹의 계통 발생학적 나무 내 위치를 나타낸다. SNP 기반 이름은 정의 SNP 돌연변이를 참조하며, R-M343은 R1b와 동일하다.[20] 계통 발생학적 이름은 새로운 발견에 따라 변경될 수 있으며,[12] SNP 기반 이름도 계통 발생학적 나무 내에서 재분류될 수 있다.
2017년 1월 30일 기준 ISOGG 트리에 따른 R1b의 기본 개요는 다음과 같다.[13]
| 2= | 2=}}}} |
인도유럽어족의 계통수와 비교하면, 사템어에 속하는 발트-슬라브어파, 알바니아어, 인도이란어파를 제외하고, 언어 계통수와 유전자 계통수가 거의 일치한다.[100]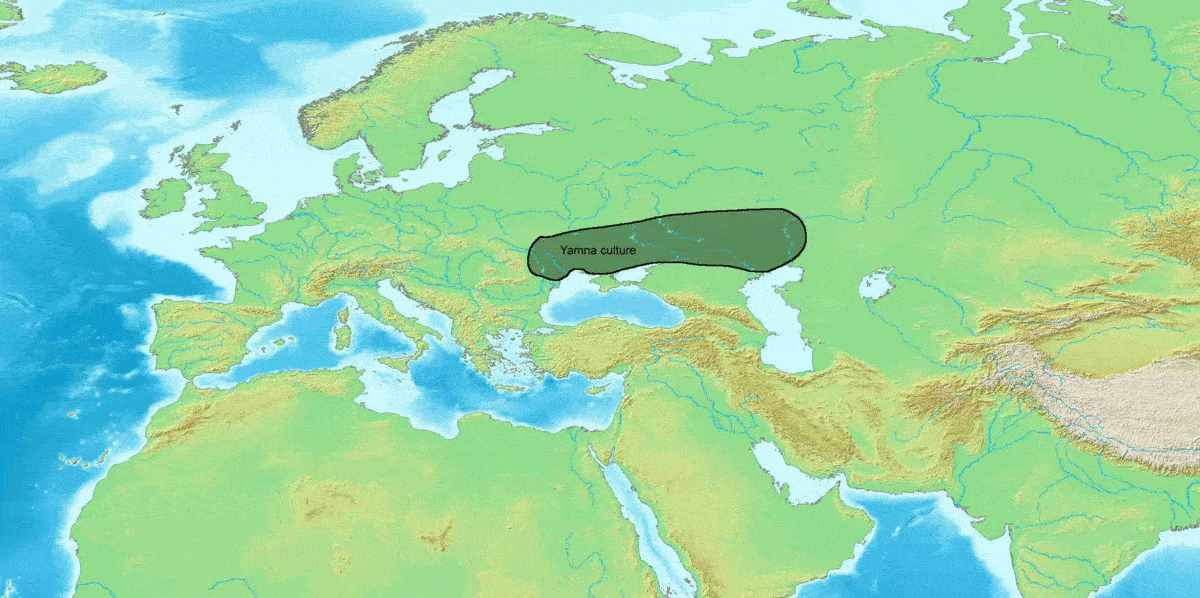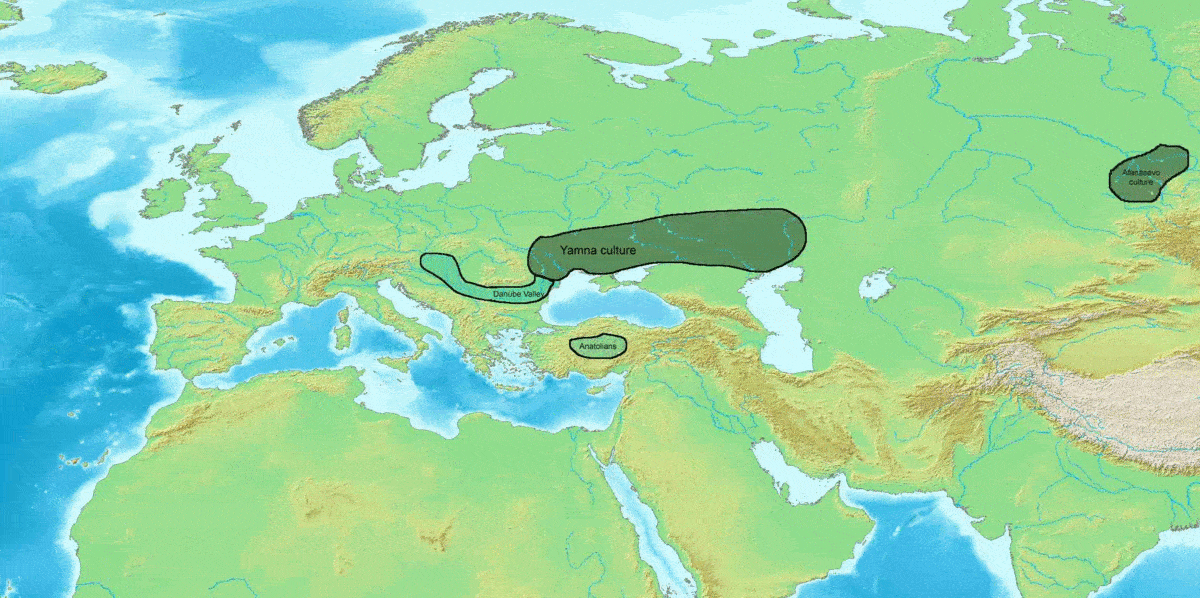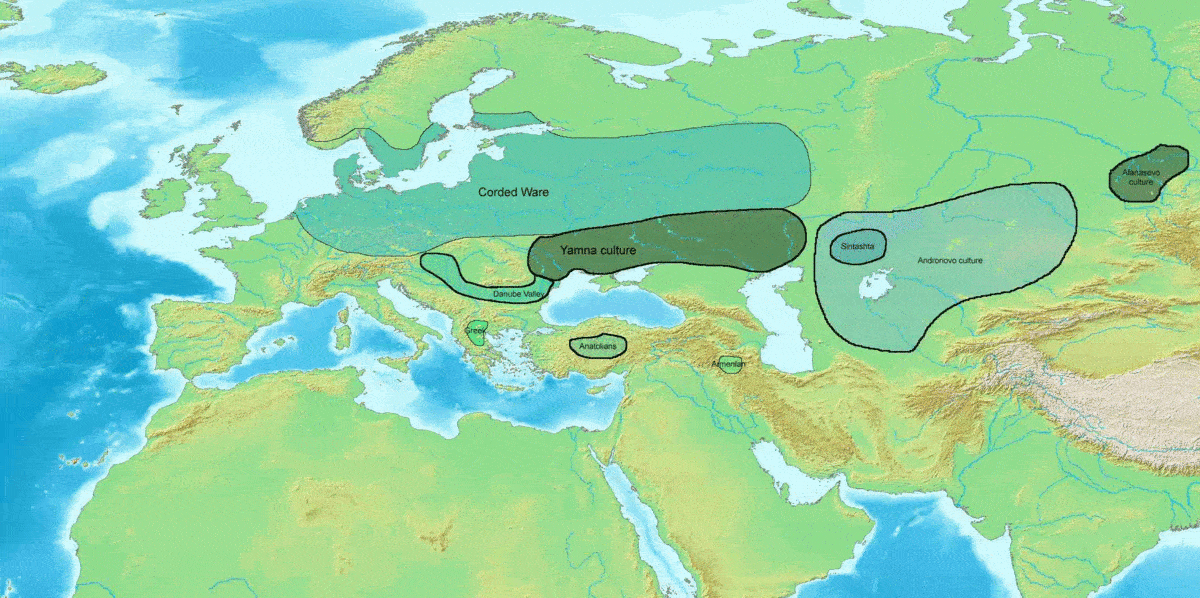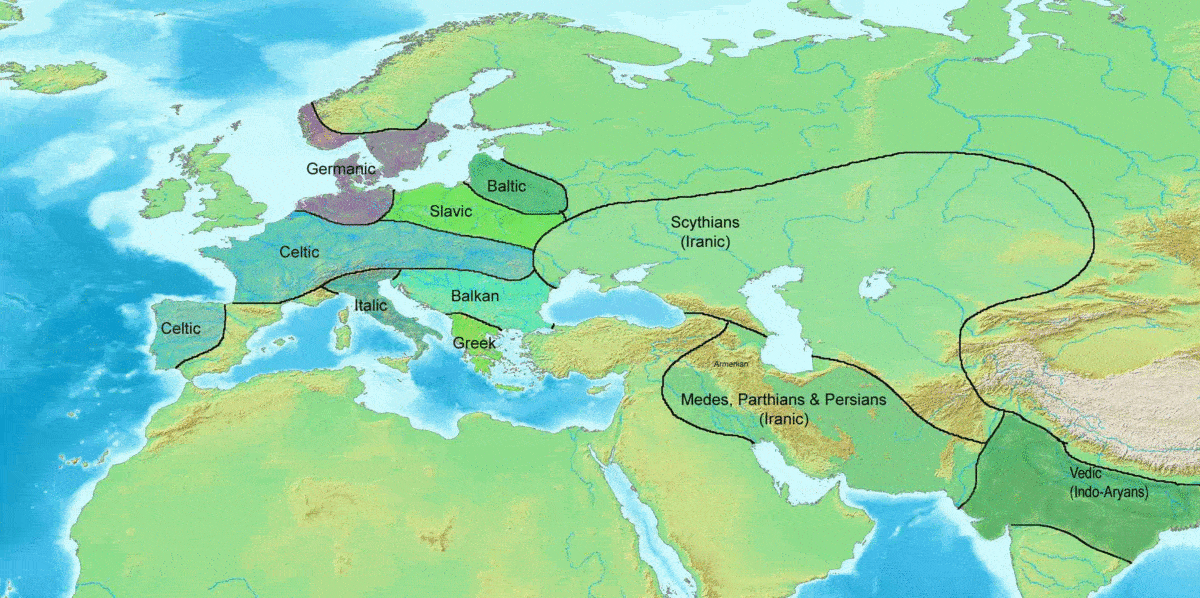
참조
[1]
논문
Population genomics of Bronze Age Eurasia
https://depot.ceon.p[...]
2015-06
[2]
논문
Eight thousand years of natural selection in Europe
[3]
논문
Neolithic and Bronze Age migration to Ireland and establishment of the insular Atlantic genome
2016-01
[4]
논문
The population genomics of archaeological transition in west Iberia: Investigation of ancient substructure using imputation and haplotype-based methods
2017-07
[5]
논문
The genetic history of Ice Age Europe
2016-06
[6]
논문
The first horse herders and the impact of early Bronze Age steppe expansions into Asia
2018-06
[7]
논문
Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia
2015-03
[8]
논문
The population history of northeastern Siberia since the Pleistocene
https://www.nature.c[...]
2019-06
[9]
논문
Ancient DNA reveals male diffusion through the Neolithic Mediterranean route
2011-06
[10]
논문
Ancient DNA from European early neolithic farmers reveals their near eastern affinities
2010-11
[11]
논문
The Beaker phenomenon and the genomic transformation of northwest Europe
2018-03
[12]
웹사이트
ISOGG 2016 Y-DNA Haplogroup R
https://isogg.org/tr[...]
[13]
웹사이트
ISOGG 2017 Y-DNA Haplogroup R
https://isogg.org/tr[...]
[14]
YFull YTree
YFull YTree v10.00.00
https://www.yfull.co[...]
[15]
웹사이트
Phylogenetic tree of haplogroup R-V1636 at 23mofang
https://www.23mofang[...]
[16]
Haplogroup
Haplogroup R on the Y-DNA Haplotree at FTDNA
https://www.familytr[...]
[17]
웹사이트
R-V1636 YTree
https://www.yfull.co[...]
[18]
논문
Ancient migratory events in the Middle East: new clues from the Y-chromosome variation of modern Iranians
2012-07-18
[19]
논문
The case of the unreliable SNP: recurrent back-mutation of Y-chromosomal marker P25 through gene conversion
https://figshare.com[...]
2006-05
[20]
논문
Excavating Y-chromosome haplotype strata in Anatolia
2004-01
[21]
논문
Human Y chromosome haplogroup R-V88: a paternal genetic record of early mid Holocene trans-Saharan connections and the spread of Chadic languages
2010-07
[22]
학위논문
Population History of the Dniester-Carpathians: evidence from Alu insertion and Y-chromosome polymorphisms
Ludwig-Maximilians-Universität München
2006
[23]
논문
Ancient links between Siberians and Native Americans revealed by subtyping the Y chromosome haplogroup Q1a
2011-08
[24]
논문
Mitochondrial DNA and Y chromosome variation provides evidence for a recent common ancestry between Native Americans and Indigenous Altaians
2012-02
[25]
논문
The genome-wide structure of the Jewish people
2010-07
[26]
논문
Polarity and temporality of high-resolution y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic influence of Central Asian pastoralists
2006-02
[27]
논문
Afghan Hindu Kush: where Eurasian sub-continent gene flows converge
[28]
논문
Human paternal and maternal demographic histories: insights from high-resolution Y chromosome and mtDNA sequences
[29]
논문
Siberian genetic diversity reveals complex origins of the Samoyedic-speaking populations
2018-11
[30]
논문
Распределение гаплогрупп И-хромосомы казахов Южно-Казахстанской, Жамбылской и Алматинской областей
http://nblib.library[...]
2021-11-20
[31]
논문
A study of genetic diversity of three isolated populations in Xinjiang using Y-SNP.
2018
[32]
문서
[33]
논문
A predominantly neolithic origin for European paternal lineages
2010-01
[34]
서적
Anthropological genetics: theory, methods and applications
Cambridge University Press
[35]
논문
Strong intra- and inter-continental differentiation revealed by Y chromosome SNPs M269, U106 and U152
2011-06
[36]
논문
Reconstructing the genetic history of Italians: new insights from a male (Y-chromosome) perspective
https://pubmed.ncbi.[...]
2018-02
[37]
학위논문
Structure of the Gene Pool of Bashkir Subpopulations
http://ftp.anrb.ru/m[...]
Institute of Biochemistry and Genetics of the Ufa Scientific Center of the Russian Academy of Sciences
2009
[38]
논문
Genetic studies on the prehispanic population buried in Punta Azul cave (El Hierro, Canary Islands)
2017
[39]
논문
Analysis of Y-chromosomal SNP haplogroups and STR haplotypes in an Algerian population sample
2008-05
[40]
논문
Genetic Testing of Language Replacement Hypothesis in Southwest Asia
[41]
논문
Haplogrouop R1b (Y-DNA)
https://www.eupedia.[...]
[42]
학위논문
Изменчивость Митохондриальной ДНК и Y-Хромосомы в Популяциях Волго-Уральского Региона
http://ibg.anrb.ru/d[...]
2015-02
[43]
논문
Neolithic patrilineal signals indicate that the Armenian plateau was repopulated by agriculturalists
2012-03
[44]
논문
Kinship and Y-chromosome analysis of 7th century human remains: novel DNA extraction and typing procedure for ancient material
2009-06
[45]
논문
The peopling of the last Green Sahara revealed by high-coverage resequencing of trans-Saharan patrilineages
2018-02
[46]
논문
Genetic history from the Middle Neolithic to present on the Mediterranean island of Sardinia
2020-02
[47]
논문
Y-chromosome and Surname Analyses for Reconstructing Past Population Structures: The Sardinian Population as a Test Case
2019
[48]
논문
Population genomics of post-glacial western Eurasia
2024
[49]
논문
Chad Genetic Diversity Reveals an African History Marked by Multiple Holocene Eurasian Migrations
https://www.cell.com[...]
2016-12
[50]
논문
Internal diversification of non-Sub-Saharan haplogroups in Sahelian populations and the spread of pastoralism beyond the Sahara
https://pubmed.ncbi.[...]
2017
[51]
논문
Population history and genetic adaptation of the Fulani nomads: inferences from genome-wide data and the lactase persistence trait
2019
[52]
논문
Northwest African Neolithic initiated by migrants from Iberia and Levant
2023
[53]
논문
Ancient genomes from North Africa evidence prehistoric migrations to the Maghreb from both the Levant and Europe
2018-06-26
[54]
논문
Isolates in a corridor of migrations: a high-resolution analysis of Y-chromosome variation in Jordan
[55]
논문
Y-chromosome variation among Sudanese: restricted gene flow, concordance with language, geography, and history
2008-11
[56]
논문
A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe
2011-01
[57]
논문
Genetic History of Chad
2018-12
[58]
논문
Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes
2005-07
[59]
논문
A back migration from Asia to sub-Saharan Africa is supported by high-resolution analysis of human Y-chromosome haplotypes
2002-05
[60]
논문
New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree
2008-05
[61]
논문
Y chromosome sequence variation and the history of human populations
2000-11
[62]
논문
Y-chromosome based evidence for pre-neolithic origin of the genetically homogeneous but diverse Sardinian population: inference for association scans
2008-01
[63]
논문
Y-chromosomal diversity in Lebanon is structured by recent historical events
2008-04
[64]
논문
Extended Y chromosome investigation suggests postglacial migrations of modern humans into East Asia via the northern route
2011-01
[65]
논문
Ladakh, India: the land of high passes and genetic heterogeneity reveals a confluence of migrations
2016-03
[66]
웹사이트
Odkud přišli Přemyslovci? Analýza DNA byla pro vědce velkým překvapením
https://www.dotyk.cz[...]
2023-05-13
[67]
논문
Genetic genealogy reveals true Y haplogroup of House of Bourbon contradicting recent identification of the presumed remains of two French Kings
2013-10-09
[68]
서적
Guardian of Ancient Egypt: Essays in Honor of Zahi Hawass
https://www.research[...]
Czech Institute of Egyptology
2020
[69]
논문
Insights from ancient DNA analysis of Egyptian human mummies: clues to disease and kinship
https://academic.oup[...]
2020
[70]
논문
The Y chromosome: a blueprint for men's health?
2017-11
[71]
논문
Limited Effect of Y Chromosome Variation on Coronary Artery Disease and Mortality in UK Biobank—Brief Report
2022-07
[72]
논문
Seeing the wood for the trees: a minimal reference phylogeny for the human Y chromosome
[73]
웹사이트
International Society of Genetic Genealogy (ISOGG; 2015), ''Y-DNA Haplogroup Tree 2015''
http://www.isogg.org[...]
2015-02-01
[74]
문서
Haplogroup A0-T is also known as A-L1085 (and previously as A0'1'2'3'4).
[75]
문서
Haplogroup A1 is also known as A1'2'3'4.
[76]
문서
F-Y27277, sometimes known as F2'4, is both the parent clade of F2 and F4 and a child of F-M89.
[77]
문서
Haplogroup LT (L298/P326) is also known as Haplogroup K1.
[78]
문서
Between 2002 and 2008, [[Haplogroup T-M184]] was known as "Haplogroup K2". That name has since been re-assigned to [[Haplogroup K2|K-M526]], the sibling of Haplogroup LT.
[79]
문서
Haplogroup K2b (M1221/P331/PF5911) is also known as Haplogroup MPS.
[80]
문서
Haplogroup K2b1 (P397/P399) is also known as Haplogroup MS, but has a broader and more complex internal structure.
[81]
문서
Haplogroup P (P295) is also klnown as K2b2.
[82]
웹사이트
K-M2313*, which as yet has no phylogenetic name, has been documented in two living individuals, who have ethnic ties to India and South East Asia. In addition, K-Y28299, which appears to be a primary branch of K-M2313, has been found in three living individuals from India. See: Poznik ''op. cit.'';
https://www.yfull.co[...]
2017-12-09
[83]
문서
Haplogroup S, as of 2017, is also known as K2b1a. (Previously the name Haplogroup S was assigned to K2b1a4.)
[84]
문서
Haplogroup M, as of 2017, is also known as K2b1b. (Previously the name Haplogroup M was assigned to K2b1d.)
[85]
웹사이트
International Society of Genetic Genealogy (ISOGG) – Y-DNA Haplogroup R and its Subclades
http://www.isogg.org[...]
[86]
논문
New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree
[87]
논문
Estimating Scandinavian and Gaelic ancestry in the male settlers of Iceland
[88]
문서
Ornella Semino et al 2000, The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective
[89]
논문
Genetic evidence for different male and female roles during cultural transitions in the British Isles
[90]
논문
The case of the unreliable SNP: recurrent back-mutation of Y-chromosomal marker P25 through gene conversion
[91]
웹사이트
Frequencies of prehistoric European mtDNA and Y-DNA from the Paleolithic to the Iron Age
http://www.eupedia.c[...]
2018-08-18
[92]
PDF
Massive migration from the steppe is a source for Indo-European languages in Europe
http://www.biorxiv.o[...]
[93]
PDF
The Population Genomics Of Archaeological Transition In West Iberia
http://www.biorxiv.o[...]
bioRxiv
[94]
뉴스
Haplogroup R1b (Y-DNA)
http://www.eupedia.c[...]
2018-08-18
[95]
뉴스
Haplogroup R1a (Y-DNA)
http://www.eupedia.c[...]
2018-08-18
[96]
문서
Lobov. Y chromosome analysis in subpopulations of Bashkirs from Russia
[97]
논문
The Genetic Heritage of the Earliest Settlers Persists Both in Indian Tribal and Caste Populations
[98]
논문
The Eurasian Heartland: A continental perspective on Y-chromosome diversity
http://www.pnas.org/[...]
[99]
논문
Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a
[100]
뉴스
Haplogroup R1b (Y-DNA)
http://www.eupedia.c[...]
2018-08-18
[101]
논문
A major Y-chromosome haplogroup R1b Holocene effect in Central and Western Europe
[102]
서적
The use of Y-chromosomal DNA variation to investigate population history: recent male spread in Asia and Europe
S.S. Papiha, R. Deka and R. Chakraborty (eds.), ''Genomic Diversity: applications in human population genetics''
[103]
간행물
Y-Chromosome lineages trace diffusion of people and languages in Southwestern Asia
[104]
웹사이트
Frequencies of prehistoric European mtDNA and Y-DNA from the Paleolithic to the Iron Age
http://www.eupedia.c[...]
2018-08-18
[105]
뉴스
Haplogroup R1b (Y-DNA)
http://www.eupedia.c[...]
2018-08-18
[106]
논문
Human Y chromosome haplogroup R-V88: a paternal genetic record of early mid Holocene trans-Saharan connections and the spread of Chadic languages
2010
[107]
웹사이트
European Prehistory, Anthropology & Genetics
http://www.eupedia.c[...]
2018-08-18
[108]
웹사이트
The Tutankhamun DNA Project
http://www.igenea.co[...]
[109]
뉴스
Half of European men share King Tut's DNA
http://uk.reuters.co[...]
U.K.
2018-08-18
[110]
서적
世界食物百科
原書房
1998
[111]
웹사이트
International Society of Genetic Genealogy (ISOGG) – Y-DNA Haplogroup R and its Subclades
http://www.isogg.org[...]
[112]
논문
The genetic history of Ice Age Europe
https://www.research[...]
2016-06-09
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com